The latitudinal tree species diversity gradient is mediated by mycorrhizal turnover
Public Proposal
active 3 weeks ago
Array Proposal
Group PIs
- Organized by
-
-
The latitudinal diversity gradient, which describes how species diversity peaks in the tropics and declines towards polar latitudes, is among the most general patterns characterizing life on earth. Recent global analyses have established that the latitudinal decline in tree species diversity coincides with a shift in the dominant form of mycorrhizal symbiosis, from arbuscular mycorrhizal (in the tropics) to ectomycorrhizal (in the temperate zone). I have compelling evidence that the latitudinal tree species diversity and mycorrhizal gradients are not just correlated, but binary expressions of a single biogeographic pattern. I wish to expand and publish existing preliminary analysis, which uses GFBI data (and requires permission from all participating GFBI members)
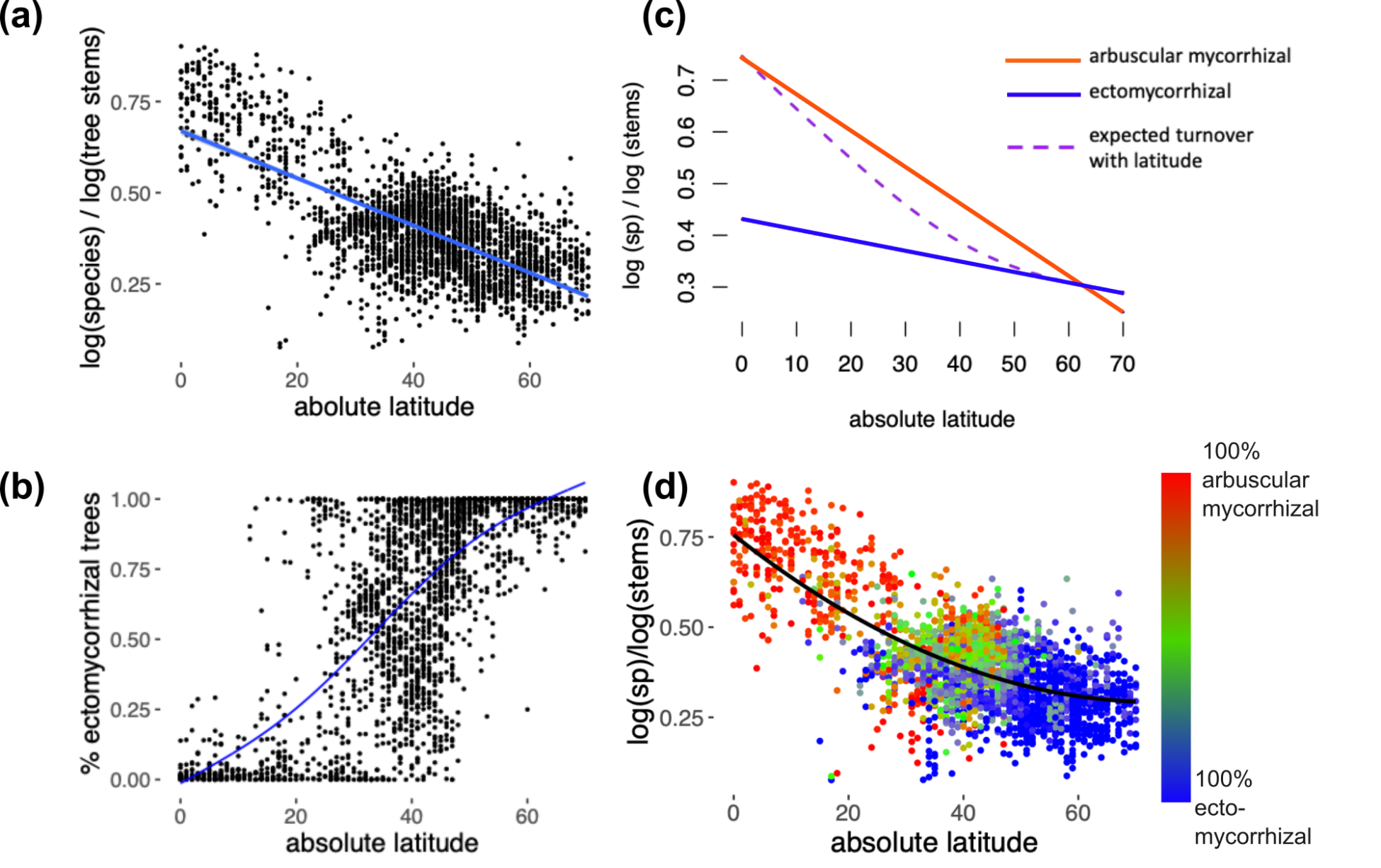
To test how independent latitudinal diversity and mycorrhizal gradients are, I used mediation analysis, which quantifies how an independent variable (latitude) influences a dependent variable (diversity) via a third variable (ectomycorrhizal abundance), known as the mediator. Specifically, I found that mycorrhizal turnover mediates up to 73% of the latitudinal diversity gradient, measured as the tree species accumulation rates (the log(tree species):log(tree stems) ratio). Another way of saying this: the majority of latitudinal effects on tree species diversity are mediated by mycorrhizal turnover.
By comparing the species accumulation curves of ecto- and arbuscular-mycorrhizal trees using a global dataset, I found that shifts to ectomycorrhizal dominance result in a decline in tree species diversity, particularly at tropical latitudes. Moreover, within a forested biome, increasing the abundance of ectomycorrhizal trees reduces the rate of species accumulation of arbuscular mycorrhizal trees. Arbuscular mycorrhizal trees have the opposite effect, increasing ectomycorrhizal tree species accumulation rates, particularly in temperate latitudes. These patterns are consistent with ectomycorrhizal trees interfering with diversity fostering mechanisms among arbuscular mycorrhizal tree species, potentially via reducing negative plant soil feedbacks.
These analyses have looked at differences in species accumulation rates without considering limitations imposed by differences in the regional species pools of the mycorrhizal groups. There are substantially more arbuscular mycorrhizal tree species than ectomycorrhizal ones. Published estimates give a figure of around 8,500 ectomycorrhizal tree species, or around 11.6% of all tree species (the remaining being overwhelmingly arbuscular mycorrhizal) (Brundrett and Tedersoo 2018). Thus, it is possible that differences in tree species accumulation rates between mycorrhizal groups reflects only differences in regional species pool sizes.
Alternatively, differences in the local ecological interactions between mycorrhizal groups can drive differences in species accumulation rates independently of species pools. Ectomycorrhizal trees generally have positive to weaker negative density dependence than arbuscular mycorrhizal ones (Bennett et al. 2017; Kadowaki et al. 2018; Liang et al. 2020; Delavaux et al. 2023). One hypothesis describing this pattern is that ectomycorrhizas sheath root tips with mycelium, making them less vulnerable to soil pathogens (Marx 1972; Laliberté et al. 2015; Bennett et al. 2017; Chen et al. 2019). Thus, all other things being equal, a system that is dominated by ectomycorrhizal trees should reduce the diversity-fostering effects of conspecific negative density dependence on ectomycorrhizal trees.
One well-developed type of analysis that can distinguish between these possibilities is a comparison of local vs. regional species richness between the two groups. This is done by comparing the plot-level potential species richness (e.g., via a Chao2 diversity estimator, which estimates the number of species under exhaustive sampling effort) with the size of the regional species for arbuscular and ectomycorrhizal trees (e.g., the total number of tree species within a WWF ecoregion). A linear (type-1) response between local and regional richness indicates proportional sampling from a regional pool (such that you would get more species in a plot if there were more species in a region). By contrast, a saturating (type-2) response indicates that local processes (e.g., competitive exclusion, niche overlap, positive density dependence) limit recruitment of species from the regional pool.
If ectomycorrhizal trees are limited primarily by their regional pools, then there should be similar proportional sampling (type-1 response) between local vs. regional species richness for ecto- and arbuscular mycorrhizal trees.
By contrast, if ecotmycorrhizal trees are more limited by local processes (e.g., weaker negative density dependence), then ectomycorrhizal trees should have either a lower slope (for type 1 response) or a saturating response (type-2).
I currently have a draft manuscript describing my existing preliminary analysis. Complementing this work with a local vs. regional species richness analysis would allow me to answer why these groups differ in their species accumulation rates — and whether it is because of the relatively young age, narrow taxonomic range of ectomycorrhizal species, or because of differences in their local ecological interactions. I consider this proposed, additional analysis indispensable for delivering a high-impact manuscript.
Finally, let me get a bit philosophical and say that the history of science has been one of describing how a diversity of phenomena can be collapsed into a small number of overarching rules. James Clerk Maxwell described how electricity and magnetism were two aspects of a single field; Einstein showed us how space and time wend together to give the fabric of the cosmos; and Darwin, most spectacularly of all, described a process that can take a single lineage of bacteria and produce all the diverse species of our biosphere.
Science has repeatedly advanced by finding unity and common cause among diversity of outcomes.
We currently have two fundamental biogeographic patterns that strikingly and reliably affect the diversity and composition of life on earth — the latitudinal diversity gradient and the latitudinal mycorrhizal gradient — that are casually linked in a way that no one has yet documented. If we can deliver a huge dataset and analysis that demonstrate this (and we absolutely can, and fast), it will be a huge advancement for the field of ecology.
And, because of the potential for high impact, I would aim for Nature, Science, or PNAS for submission.
Referenced Here:
Bennett, J. A., H. Maherali, K. O. Reinhart, Y. Lekberg, M. M. Hart, and J. Klironomos. 2017. Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science 355:181–184.
Brundrett, M. C., and L. Tedersoo. 2018. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytologist 220:1108–1115.
Chen, L., N. G. Swenson, N. Ji, X. Mi, H. Ren, L. Guo, and K. Ma. 2019. Differential soil fungus accumulation and density dependence of trees in a subtropical forest. Science 366:124–128.
Delavaux, C. S., J. A. LaManna, J. A. Myers, R. P. Phillips, S. Aguilar, D. Allen, A. Alonso, et al. 2023. Mycorrhizal feedbacks influence global forest structure and diversity. Communications Biology 6:1–11.
Kadowaki, K., S. Yamamoto, H. Sato, A. S. Tanabe, A. Hidaka, and H. Toju. 2018. Mycorrhizal fungi mediate the direction and strength of plant–soil feedbacks differently between arbuscular mycorrhizal and ectomycorrhizal communities. Communications Biology 1:1–11.
Laliberté, E., H. Lambers, T. I. Burgess, and S. J. Wright. 2015. Phosphorus limitation, soil-borne pathogens and the coexistence of plant species in hyperdiverse forests and shrublands. New Phytologist 206:507–521.
Liang, M., D. Johnson, D. F. R. P. Burslem, S. Yu, M. Fang, J. D. Taylor, A. F. S. Taylor, et al. 2020. Soil fungal networks maintain local dominance of ectomycorrhizal trees. Nature Communications 11:2636.
Marx, D. H. 1972. Ectomycorrhizae as Biological Deterrents to Pathogenic Root Infections. Annual Review of Phytopathology 10:429–454.
Steidinger, B. S., T. W. Crowther, J. Liang, M. E. V. Nuland, G. D. A. Werner, P. B. Reich, G. Nabuurs, et al. 2019. Climatic controls of decomposition drive the global biogeography of forest-tree symbioses. Nature 569:404.